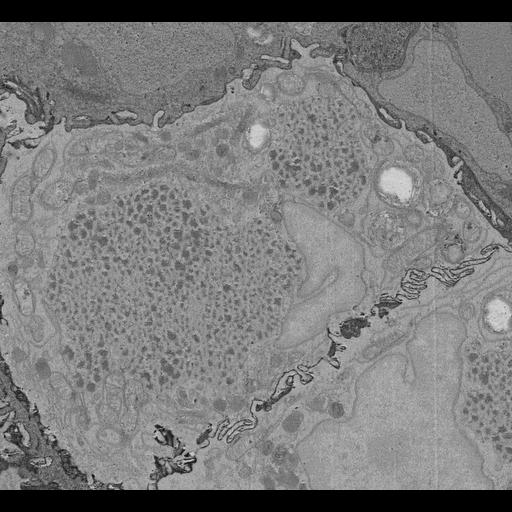
In this study we employed a voltage sensitive, positron emission tomography (PET) tracer known as fluorobenzyl triphenylphosphonium (FBnTP) (Momcilovic et al., (2019) Nature), to profile mitochondrial bioenergetics in autochthonous mouse models of lung cancer. By coupling the glucose analog PET tracer, fluoro-2-deoxy-D-glucose (F-FDG), we identified two distinct tumor sub-populations – lung adenocarcinomas (LUAD), characterized by FBnTPHI and F-FDGLO uptake and lung squamous cell carcinomas (LUSC), characterized by FBnTPLO and F-FDGHI uptake. The structural organization of the mitochondria directly impacts the bioenergetic capacity of both normal and tumor cells. We therefore explored the architecture of mitochondria in in vivo lung tumors with different PET signatures using 3D serial block-face electron microscopy (SBEM) in order to understand how mitochondrial networks impact cellular respiration and metabolism. Pairing PET imaging with 3D SBEM enabled us to functionally image mitochondria activity in whole lung tumors followed by mapping of mitochondrial ultrastructures down to the resolution of cristae. Our ultrastructure analysis of mitochondrial networks in LUAD and LUSC identified distinct mitochondrial subpopulations exist that carry equally distinct bioenergetic phenotypes. We discovered that mitochondrial networks in LUADs were distinguished by the predominance of lipid droplet bound mitochondrial subpopulations that support cellular respiration. In contrast, we identified in LUSCs that glucose flux is a key regulator of cristae remodeling and the cellular localization of mitochondrial networks. Structure-and-function studies defining the relationship between mitochondrial architecture and metabolic dependencies may hold promise as strategies to profile metabolic liabilities unique to lung cancer subtypes. We anticipate that coupling PET imaging with 3D SBEM will have dynamic applications beyond that of lung cancer and enrich our understanding of how mitochondrial bioenergetics impact human disease.
Post-fixed tissues of mouse lung tumor were washed in 0.15M cacodylate buffer (2mM Ca2+). Tissues were stained for 1 hour with 2% osmium and 1.5% potassium ferrocyanide in 0.15M cacodylate buffer (2mM Ca2+). Tissues were washed 5 times (5 minutes each time) with ddH2O, then Placed in filtered TCH buffer (0.05g thiocarbohydrazide in 10mL ddH2O) for 20 minutes at room temperature (RT). Tissues were washed 5 times (5 minutes each time) with ddH2O and then stained with 2% osmium in ddH2O for 30 minutes at RT. Following that, tissues were stained with 2% uranyl acetate overnight at RT and lead aspartate solution (0.66% (w/v) lead in 0.03M aspartic acid) for 30 minutes in 60℃ oven. Tissues were dehydrated in serial ice-cold ethanol (70%, 90% and 100%) and ice-cold acetone after washing in ddH2O. Tissues were then embedded in serial Durcupan resin (50%, 75%, 100%) and solidified in 60℃ oven for 2 days.